Introduction
DNA barcoding is a simple and useful step toward understanding the ecosystem. It also serves to further our interests in biodiversity research [1]. A short standardized sequence (400-800 bp) of DNA can be used to distinguish individuals of a species. This approach was taken, because genetic diversity between species is markedly greater than that within species [2]. Numerous computational analysis methods and systems have been introduced for this purpose [3-5]. The use of this system can provide rapid, accurate, cost-effective, and automatable process for species identification. The success rate of each barcoding application varies significantly among groups. Moreover, global datasets that represent extensive ecosystems are expected to be subjected to particular difficulties, especially in groups in which recent speciation rates are high and effective population sizes are large and reasonably stationary [6]. Several studies of species-level identification have covered many groups of organisms, including birds, fishes, and various arthropods [4, 6-8].
In order to use the barcoding system for species identification, cytochrome c oxidase subunit I (COI) sequences were obtained in this study from 529 sequences, representing 156 species from fish, insects, and shellfish in the Korean peninsula.
Methods
The first community-level barcoding studies were conducted in the most diverse terrestrial and marine ecosystems in an inland and coastal area of South Korea (include reference). We collected samples to obtain an overview of the variation patterns for 529 COI sequences among 68 fish species, 29 insect species, and 59 shellfish species. Multiple specimens were collected for most of the species. Fish and shellfish were collected from Yeosu in Jeollanam-do; shellfish were collected from Taean; and insects were collected from Chungcheongnam-do, Gangwon-do, Gyeongsangbuk-do, and Jeollabuk-do in South Korea. Samples were collected using different, technically appropriate methods (Fig. 1, Supplementary Table 1) [9]. If possible, the samples were obtained from widely distributed places in South Korea.
Genomic DNA was isolated from samples using the Qiagen DNeasy 96 blood and tissue kit (Qiagen, Valencia, CA, USA) according to the instructions. DNA fragments of target genes were amplified by polymerase chain reaction (PCR) with primers for the COI gene (primer sequences: LCO1490 GGTCAACAAATCATAAAGATATTGG and HCO 2198 TAAACTTCAGGGTGACCAAAAAATCA) [10]. PCR amplification was performed using Top-Taq PreMix (2×; CoreBio, Seoul, Korea) under the following conditions: denaturation (1 min at 94℃), annealing at 51℃ for amplification of the COI gene, and extension (2 min at 72℃). PCR products were purified with the Core-One PCR purification kit (CoreBio), and TA cloning was performed using the pGEM-T Easy Vector system (Promega, Madison, WI, USA) by Macrogen Inc. The clones for each marker were sequenced with forward (SP6) and reverse (T7) primers using an ABI 3730XL sequencer (Applied Biosystems, Foster City, CA, USA). The sequences reported in this paper have been deposited in GenBank under accession numbers HM180413-HM180941.
To obtain the species information for each operational taxonomic unit (OTU) in a phylogenetic tree, a BLAST search was performed using the BLASTN program from NCBI [11]. A cutoff value for the BLAST result was established as follows: query coverage > 90% and identity > 75% for COI. The levels of sequence divergence within and between the selected species were investigated using the pairwise Kimura 2 parameter (K2P) distance model [12]. The neighbor-joining tree, with gap positions ignored on a pairwise basis, was constructed using the neighbor-joining (NJ) method with K2P distances in MEGA4 [13]. These distances were hierarchically arranged in accordance with intraspecific and interspecific species differences within each genus. When the sequence dataset consisted of only 2 genera from the same family, an intergeneric comparison within the family was not performed.
Results and Discussion
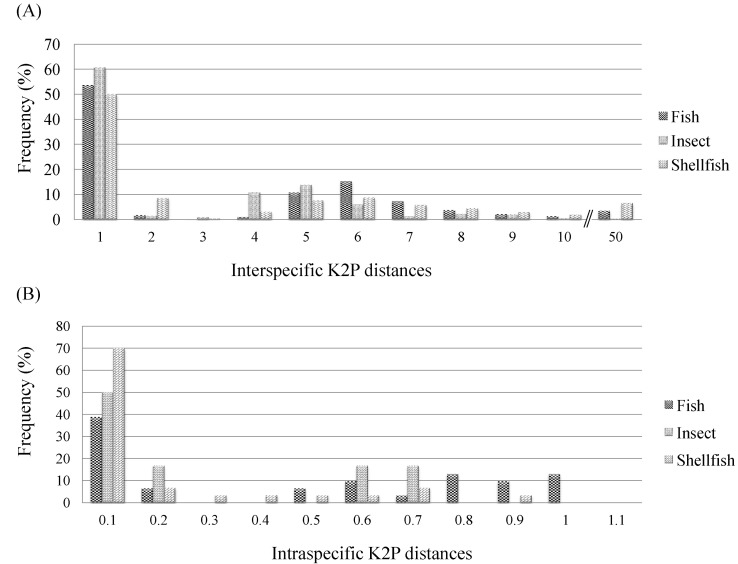
After BLASTN annotation analyses were conducted, K2P distances were compared at different taxonomic levels, revealing distinct features in the sequences both within and between species. With respect to the COI sequences of the 156 species represented, the interspecific K2P distances for the COI sequences from the 68 fish species, the 59 shellfish species, and the 29 insect species ranged from 0% to 45.25% (fish, 0% to 40.99%; insects, 0% to 10.34%; shellfish, 0% to 45.25%) (Fig. 2A), whereas the intraspecific K2P distances with ≥3 sequences ranged from 0% to 0.985% (fish, 0% to 0.985%; insects, 0.005% to 0.635%; shellfish, 0% to 0.817%) (Fig. 2B). The average interspecific distances and average intraspecific distances were, respectively, 3.58 and 0.14 in shellfish, 3.22 and 0.41 in fish, and 2.06 and 0.25 in insects (Table 1). In shellfish, the greatest interspecific K2P differences were 25.57-fold higher than the intraspecific values. The overall base composition in each species of fish, insect, and shellfish was as follows: T (thymine) ranged from 27.4% to 33.7% (highly abundant); G (guanine) ranged from 16.8% to 21.5% (not highly abundant) (Table 1). These findings for fish were consistent with previous studies showing that T occurred more frequently and G occurred less frequently than A (adenine) and C (cytosine) [8].
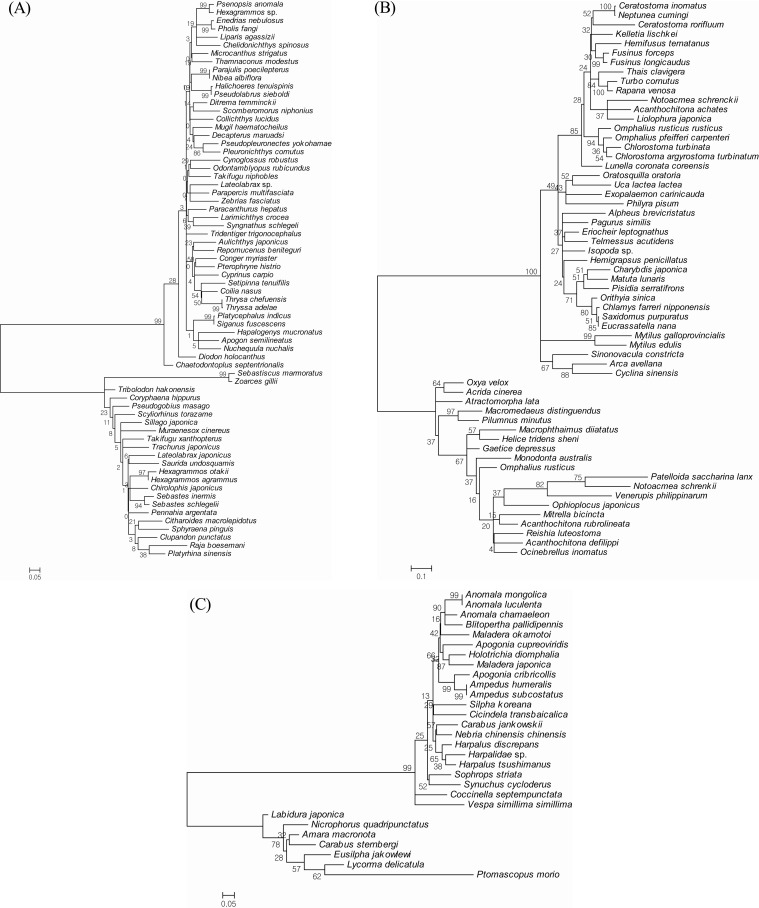
In our polytypic species analysis with more than 3 individuals in each species, the average intraspecific difference was approximately 0.5%, and the maximum intraspecific divergence was only 1.86% (Table 2). The highest overall GC% content was found in the 18 species of fish. Lower values were found in the 2 species of insects and in the 6 species of shellfish (Table 2). The fish Chelidonichthys spinosus had a high GC% content of 50.9%. The mean GC% content of the 18 barcoded fish species was higher than that of the 6 shellfish species (46.9 ± 2.2% vs. 38.0 ± 4.9%) (see also Table 2). Sixteen of the 21 species with GC% content ≥45% were fish, whereas only 1 shellfish species exhibited GC% content ≥45%. The GC% content can be used in a new approach to evaluate animal evolutionary relationships, although the relationship between GC% content and the evolutionary branching date is not very accurate [14]. Moreover, the average divergence of congeneric species pairs was greater than that found for intraspecific differences, but 10 species in 5 genera had interspecific distances below 0.1% (Table 3). These species included Hexagrammos agrammus/H. otakii, Ampedus humeralis/A. subcostatus, Anomala luculenta/A. mongolica, Chlorostoma argyrostoma turbinatum/C. turbinate, and Omphalius rusticus rusticus/O. pfeifferi carpenteri. In addition, the NJ tree exhibited shallow interspecific divergence except at the first deep divergence (Fig. 3). In fish, several clades had a high level of bootstrap support (≥97%) (Fig. 3A). These clades included Thrysa chefuensis and T. adelae, Hexagrammos otakii and H. agrammus. In insects, the clades that had a high level of bootstrap support (≥95%) included Fusinus forceps, F. longicaudus, Mytilus galloprovincialis, and M. edulis. In shellfish, 2 clades separated out with a high level of bootstrap support (≥99%) (Fig. 3B). These clades included Anomala mongolica and A. luculenta, Ampedus humeralis and A. subcostatus (Fig. 3C).
In conclusion, we obtained DNA barcodes using COI sequences from fish, insects, and shellfish. The aims of this research were species identification and contribution to biodiversity research. At the species level, the rate of correct identifications might be low in a diversified environment. However, DNA barcoded sequences can be used for the interpretation of species-level identification and community-level patterns in fish, insects, and shellfish.