Introduction
Breast cancer (BC) is the most common cancer in females worldwide, with nearly 2.3 million new cases diagnosed in 2020. It accounts for approximately 11.7% of all cancer cases and 24.5% of all cancer cases in females [1]. Since 2008, the global incidence of BC has increased by more than 20%, and the death rate has risen by 14% [2]. As a result, research on relevant tumor indicators for early diagnosis and monitoring is required, and recent studies have concentrated on the role of the immune system in cancer progression [3].
BC is a multi-step process that involves numerous genetic changes, such as oncogene activation and cancer suppressor gene inactivation [4]. The gene encoding interleukin (IL)-18 is found on chromosome 11 at positions 11q22.2âq22.3 and has six exons in humans. IL-18 is a multidirectional cytokine that regulates the immune response in various ways; in 1989, IL-18 was first described as an âinterferon-inducing factor.â IL-18 plays an essential role in stimulating natural killer cells, and cellular anticancer activities also enhance the expansion of Th1 and cell activation. Furthermore, IL-18 increases the expression of adhesion-related molecules, nitric oxide synthase enzyme synthesis, and chemokine production [5]. IL-18, in combination with IL-2, causes a Th2 cell response and the production of IL-4 and IL-13. Simultaneously, IL-18 can reduce antitumor immunity in a programmed-death-1 (PD-1) dependent manner. PD-1 is a co-inhibitory receptor that constitutes one of the top checkpoints. Many polymorphisms in the IL-18 promoter region affect transcript factor binding locations [6], which could be IL-18 expression quantitative trait loci. Several studies have found that cytokine gene polymorphisms impact cytokine production, which may be linked to disease [7].
Single-nucleotide polymorphisms (SNPs) are found in the IL18 gene, particularly in the promoter region bound to â607G/T (rs1946518) and â137G/C (rs187238). These SNPs (â137G/C and â607G/T) in the IL18 gene promoter region are expected to affect IL-18 expression and activity [8]. Arimitsu et al. [9] found that monocytes in individuals with the 137G/G genotype produced considerably more IL-18 than those in individuals with the 137G/C genotype. Furthermore, a link between these two SNPs and cancer susceptibility has been established [10]. Research on the relevance of IL18 polymorphisms in BC risk remains contentious. To our knowledge, only three studies have examined the influence of IL18 genetic variations (â137G/C and â607A/C) on BC susceptibility; Khalili-Azad et al. [11] studied the effect of IL18 polymorphisms (â607A/C and â137G/C) on BC risk in 200 patients and 206 healthy controls. They discovered that CC homozygosity for the â137G/C polymorphism lowered the risk of BC [11]. Other studies found that the IL18 â607A/C polymorphism contributed to an increased risk of BC [12,13]. In light of this information, it is necessary to investigate IL18 SNPs in Iraqi women with BC.
Methods
After the ethical committee of the Medical City directorate approved this study, 100 women who visited the oncology teaching hospital's breast clinic were recruited from January 28, 2020 to August 11, 2022. Group 1 included 60 women who had recently been histopathologically diagnosed with BC and provided written informed consent for participation in the study. Group 2 included 40 healthy women who served as controls. Patients with other systemic diseases and those taking any hormone-modifying drug were excluded from the study.
DNA extraction and genotyping
Each patient and control had 8-10 mL of blood taken. Blood samples were taken from the cubital vein and placed directly into an anticoagulant tube containing EDTA. Genomic DNA extraction was performed using a Norgen Biotek kit (Thorold, ON, Canada). The optical density ratio at 260/280 nm was used to assess the quantity and quality of isolated DNA Using a Nano-Drop device (Qubit 4, Invitrogen, Waltham, MA, USA). It was preferable to have a balance of 1.7â1.9. Electrophoresis on a 1% agarose gel was performed to corroborate the findings. Until the genotyping analysis, the DNA samples were stored at â20°C.
Polymerase chain reaction
Polymorphisms were investigated by utilizing sequence-specific amplification-polymerase chain reaction (SSP-PCR) at locations â607C/A rs1946518 and â137G/C rs187238 within the promoter region of IL18. For the targeted location â607C/A, a common reverse primer and two specific forward primers were utilized, with an amplified product size of 196 bp, and a forward control primer was used to amplify a 301-bp fragment covering the polymorphism region as an internal control (Table 1).
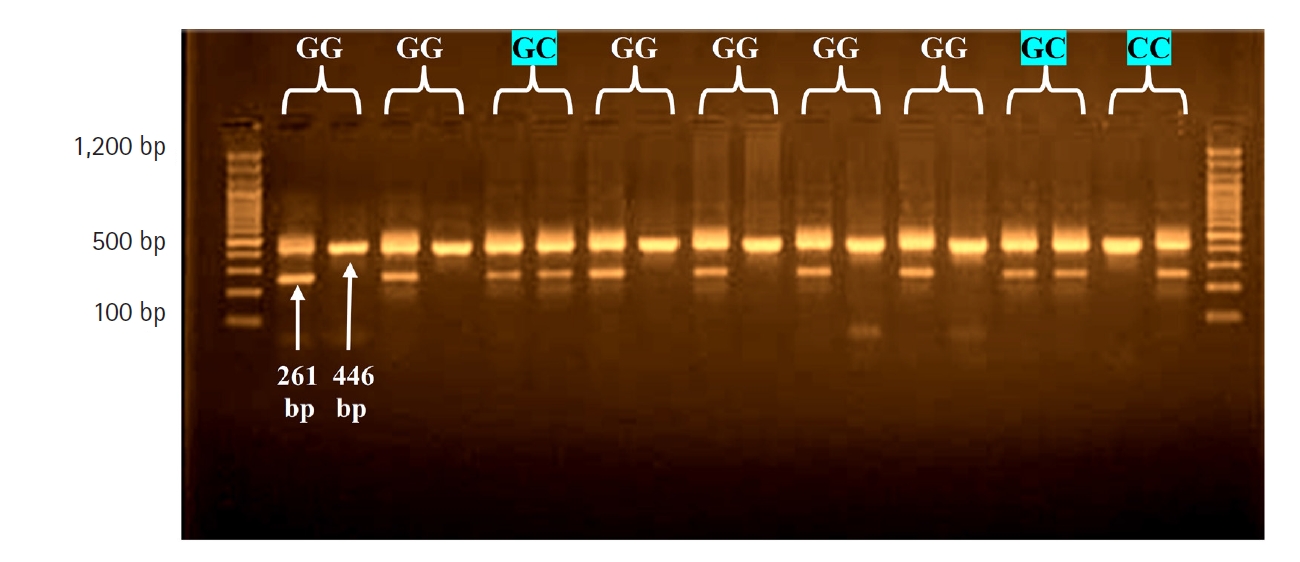
In the location â137G/C, a common reverse primer and two specific forward primers were also utilized, with an amplified product size of 261 bp, and a forward control primer was used to amplify a 446-bp fragment covering the polymorphism region as an internal control (Table 2). The polymerase chain reaction (PCR) reactions were carried out in a volume of 25 ÎźL, including 12.5 ÎźL of 2Ă Go-Taq Green Master mix (Promega, Madison, WI, USA), 3 ÎźL of genomic DNA, and 6.5 ÎźL of nuclease-free water. All reaction mixtures contained one sequence-specific primer, one common reverse primer, and one internal position control primer, each at a concentration of 1 ÎźL. As a result, each piece of DNA was subjected to two PCR tests: one for the F1 wild-type allele and one for the F2 mutant allele [8]. The assays were carried out in a thermocycler (Applied Biosystems, Waltham, MA, USA). Denaturation was carried out at 95°C for 5 min, followed by 30 cycles at 94°C for 30 s, 57°C for 30 s, 72°C for 40 s, and 72°C for 10 min. Then, the PCR results were visualized using electrophoresis on 2% agarose gel, and the molecular weight was calculated using a 100-bp DNA ladder.
Statistical analysis
The genotype of cytokines was statistically analyzed using the SPSS version 25 (IBM Corp., Armonk, NY, USA). The allele frequencies of the cytokine genes were calculated by the direct gene counting method. A freely available online calculator was used to calculate whether there was a significant departure from Hardy-Weinberg (H-W) equilibrium for two alleles (http://www.had2know.com/academics/Hardy-Weinberg equilibrium-calculator-3-alleles.html). H-W equilibrium is the expected frequency of genotypes if mating is non-assortative and there are no mutations from one allele to another. When there are two alleles for a particular gene, A and B, and their respective population frequencies are p and q, the expected frequencies of the genotypes AA, AB, and BB are p2, 2pq, and q2, respectively. The Pearson chi-square test was used to assess whether there were significant differences between the observed and expected frequencies. The alleles and genotypes of cytokines were presented as percentages and frequencies, and the two-tailed Fisher exact test was calculated to assess the significance of differences between their distributions in BC patients and controls. Odds ratios (ORs) were also estimated to define the association between cytokine alleles and genotypes with BC. OR values can range from between 0 and 1 (for a negative association) to more than 1 (for a positive association).
Results
IL-18 â607C/A (rs1946518) SSP-PCR
This study analyzed the distribution of the genotype and allele frequencies of the rs1946518 polymorphism (at the â607C/A position) in patients and controls (Fig. 1). This polymorphism presented three genotypes (CC, CA, and AA) that corresponded to two alleles (T and A) in BC patients and controls. The genotype frequencies in both groups were not in agreement with H-W equilibrium, and there were highly significant differences between the observed and expected frequencies in both patients and controls (Ď2 = 3.16 and Ď2 = 16.5), respectively (Table 3).
The CC genotype was significantly less common in patients than in controls (15.0% vs. 65.5%, p < 0.001), and the magnitude of this negative association was 0.11. However, the AA genotype frequency was higher in patients (51.7%) than in controls (22.5%); this difference was significant (OR, 2.83; p = 0.004). The CA genotype frequency was also significantly higher in patients than in controls (33.3% vs. 15.0%; OR, 3.68; p = 0.04). The C allele frequency was lower in patients than in controls (25.0% vs. 38.7%), while the A allele frequency was higher (75.0% vs. 61.3%, respectively). A negative association was found for the C allele (OR, 0.19) and a positive association for the A allele (OR, 5.03) (p < 0.001 for both) (Table 4).
IL-18â137G/C (rs187238) SSP-PCR
The rs187238 genetic polymorphism of the IL18 gene was investigated at position â137G/C (Fig. 2), and presented with three genotypes (GG, GC, and CC) that corresponded to two alleles (G and C). The results of patients were in agreement with the expected H-W equilibrium. However, in the control group, the results were not in agreement with the expected (H-W) equilibrium, and there was a highly significant difference between the observed and expected frequencies (Ď2 = 5.74) (Table 5).
At the position â137G/C in IL18, patients had a significantly higher frequency of the GC genotype than controls (43.3% vs. 22.5%; OR, 2.63; p = 0.04). In contrast, the frequency of the CC genotype was significantly higher in patients than in controls (23.3% and 12.5%; OR, 2.13). Therefore, the G allele frequency was significantly lower in patients than in controls (55.0% vs. 76.2%, respectively), with a highly significant difference (p = 0.002). A negative association was found for the G allele (OR, 0.38) and a positive association for the C allele (OR, 2.36) (Table 6).
Discussion
The present study investigated the two most commonly studied SNPs of the IL18 gene (â607C/A and â137G/C). The patients and controls both showed deviation from H-W equilibrium for the -607C/A genotype, which may have been related to BC or intermarriage in Arab Iraqi society between relatives. This result aligns with that of a previous study on BC [14].
Although the distribution of the â137G/C genotype in the patient group was consistent with H-W equilibrium, this was not the case for the control group, which showed a highly significant difference between the observed and expected frequencies (Ď2 = 5.74). This SNP may be considered a common genotype in the Iraqi population, with the wild-type G allele having a protective function, reducing susceptibility, and the mutant C allele having an environmental effect.
These SNPs' genotypes and alleles showed significant differences between BC patients and controls. The present study observed that the IL18 â607 CA and AA genotypes were present in about 85% of BC cases. The wild-type CC genotype had a low frequency (about 15%) in BC patients, which may highlight the role of the IL18 â607 polymorphism in the pathogenesis of disease. The A allele was an environmental effect allele, while the C allele had a preventive fraction because the CC genotype showed the highest frequency in the control group (70%). These highly significant findings for the AA and CA genotypes at position IL18 â607 suggest that this polymorphism may play a role in cancer progression.
Several studies on IL18 polymorphisms have been conducted in various populations in multiple countries; one of them has found a link between IL18 polymorphisms and the risk of BC [15]. The IL18 â607C/A polymorphism may be linked to an increased risk of BC in Asian and mixed populations [16]. Furthermore, the present results showed that the IL18 rs1946518 SNP might play a role in BC because the wild-type allele C of the â607 SNP had a protective effect against BC (OR, 0.19). In contrast, the mutant allele A had a positive association (OR, 5.03) suggesting an etiological impact; therefore, women who carry allele A of â607 may be more susceptible to BC than women who have allele C. However, the results of some studies were mixed; in a study involving 72 BC patients and 93 control women, Fathi Maroufi et al. [17] discovered that the IL18 -607A/C polymorphism was not linked to BC in an Iranian population sample. The IL18 rs187238 polymorphism results showed no significant differences according to the CC genotype frequencies between BC patients and controls. Nonetheless, it was interesting to note in the present study that the heterozygous GC genotype had an OR of 2.63 for the patient group, implying that the mutant allele C may have had an environmental effect on the Iraqi population, conferring susceptibility to BC, while the wild-type allele G had a preventive effect against BC (OR, 0.38). Our findings show that the IL18 -137G/C polymorphism is associated with the development of BC. The conversion of G (guanine) to C (cytosine) at position -137G/C of the IL18 gene removes a nuclear factor binding site for histone-4 transcriptional factor-1 [18]. Genetic variants have been considered the most critical cancer risk factors. Although high-penetrant capability genes (e.g., BRCA1 and BRCA2) have strong links to BC, low-penetrant susceptibility genes that predispose individuals to the disease have yet to be identified; nonetheless, immune responses and surveillance may be affected by genetic variability in a sequence of immune regulatory genes [19]. The IL18 promoter polymorphism â137G/C has previously been linked to various cancers in different populations, including esophageal squamous cell malignant tumors, prostate cancer in the Chinese population [20], colorectal cancer in Greek people [21], and ovarian cancer in native Hawaiians [22]. Additional case-control studies on BC and gastric cancer progression have been published [23,24]. However, there is no link between type 2 diabetes mellitus development and the IL18 â137G/C gene polymorphism [25].