Introduction
Cancer has a complex pathophysiology unlike other diseases. The development of cancer is attributed to a number of predisposing factors like familial and genetic factors, racial and geographic factors and various epidemiological factors like smoking, use of tobacco, diet, and certain type of infection. Majority of the cancers occur in ageing population [1], however there is a changing trend towards younger population in recent years. The uncontrolled division of cancer cells generally spreads to the other part of body through the blood and lymphatic systems. In this context, solid cancer which is an abnormal mass of tissue that usually does not contain cysts or liquid area can be either benign or malignant in nature. Such solid cancers are observed in tissues of epithelial or endothelial cells of lung, breast, prostate, pancreatic, colorectal, stomach, and bladder. Management of cancer is a multimodality approach and includes surgery, radiotherapy, chemotherapy and targeted therapy with certain limitations of each approach. Research is going across the globe for the management of cancer. Recently microRNAs (miRNAs) have drawn attention of the researchers to discover new insights into the treatment of cancer. These miRNAs are highly conserved non-coding single stranded RNAs (~18 to 22 bp) that target mRNA at 3' un-translated region of protein coding genes and act as natural inhibitor. These miRNAs participate in several biological processes such as cell differentiation, proliferation, metabolism, and apoptosis and are also involved in the regulation of gene expression [2]. However, in development of pathologies like cancer, deregulation of miRNAs expression has been identified as potential area to be studied. The miRNAs have special signature and variation from normal cells [3]. Hence, use of specific miRNAs as biomarker may be applied for cancer diagnosis and treatment. Since the expression of tumor suppressor genes and oncogenes are influenced by the miRNAs, provides scope to study their association in cancer [3]. Collating data for miRNAs signatures associated with solid tumors, it was reported that more than 50% of miRNAs genes are located in cancer associated genomic regions or in fragile sites, suggesting that miRNAs play a crucial role in the pathogenesis of a limited range of human cancers than previously expected [4]. Considering their influence on the cancer cell phenotype, some miRNAs are considered to be oncogenic, and others to be tumor-suppressive [5]. Few miRNAs are well characterized in cancer association such as miR-21, miR-17-5p, miR-191, miR-29b-2, miR-223, miR-128b, miR-199a-1, miR-24-1, miR-24-2, miR-146, miR-155, miR-181b-1, miR-20a, miR-107, miR-32, miR-92-2, miR-214, miR-30c, miR-25, miR-221, miR-106a, miR-126, and miR-31 (Supplementary Table 1) that are involved in either up regulation or down regulation of many genes [42,43,44,45,46,47,48,49,50,51,52,53,54,55,56,57,58]. Till date various researchers have explored the unusual expression of miRNAs, and its prognostic significance in various human malignancies. Hence, to study the relationship between miRNAs and cancer, it is important to examine the expression profile of miRNAs and their target genes in different cancer types. As a case study, the unusual expression of miRNA-21 has been conformed in various cancers such as lung, breast, prostate, colorectal, pancreatic, bladder, and stomach cancer, and interestingly the oncogenic like function of miRNA-21 is mediated through the expression of its target genes like PDCD4, PTEN, and TGFBR2 which are generally tumor suppressor genes in nature [59]. Though miRNAs control the gene expression to some extent during translation process but the translational suppression mechanism is still controversial, for example some miRNAs down-regulate while others up-regulate the gene expression. Hence, study of interaction mechanism between miRNAs and their target genes is essential. In the process of miRNAs mediated gene regulation, the Argonaute (AGO) class of proteins play a significant role in the hybrid formation between miRNA-mRNA [60,61]. Despite of several researches, controlling this abnormal cell proliferation selectively in different cancer by miRNAs as natural inhibitor is still a major challenge. In the present study, attempts have been made to develop a strategy for identification and selection of suitable miRNAs, their target genes associated in solid tumor, and as well as to study the role of AGO protein in their interaction at molecular level using computational tools and techniques.
Methods
Screening of miRNAs and genes
Extensive literature search and exploration of online databases were conducted to understand miRNAs mediated regulation of solid cancer associated target genes using miRCancer (http://mircancer.ecu.edu/) and miRDisease (http://mir2disease.org). miRNAs and target genes were screened by following two criteria: (1) only up regulated oncogenes or down regulated tumor suppressor genes were selected and (2) the screening was performed by selecting oncogenes and tumor suppressor genes as target genes when both of them are either up regulated or down regulated. Further, minimum free energy (MFE) score based binding affinity between miRNAs and their target genes were explored and inspected using miRTarbase web server (http://mirtarbase.mbc.nctu.edu.tw/). Furthermore, the involvement of target genes in different biological function was inspected using UniProt (http://www.uniprot.org/) web server and plotted using Venn (http://bioinformatics.psb.ugent.be/webtools/Venn/) diagram.
Prediction of structural model of miRNA-mRNA complexes
To verify the folding affinity between selected miRNAs and their target genes, secondary structures were predicted using RNAfold web server (http://rna.tbi.univie.ac.at/cgi-bin/RNAfold.cgi). The resulted dot bracketed structures of duplex were used for prediction of their tertiary structure using RNA COMPOSER (http://rnacomposer.cs.put poznan.pl/).
Structure extraction of AGO protein
The three-dimensional structure of AGO protein was retrieved from Protein Data Bank (PDB ID: 3F73). The structure preparation and correction of AGO protein were performed using Discovery Studio 3.5 suite (http://accelrys.com/products/discovery-studio/visualizatation-download.php).
Molecular docking between miRNA-mRNA and AGO protein
Docking between three dimensional structure of protein and ligand, may be a biomolecule like miRNA or a gene is an efficient computational method to inspect the molecular interaction [61]. Docking were performed between AGO protein and miRNA-mRNA duplexes (miR-106 and PTEN; miR-21 and TGFBR2; and miR-29b-2 and VEGFA) using PatchDock web server (http://bioinfo3d.cs.tau.ac.il/PatchDock/),algorithm ranked the docked complexes on the basis of highest geometrical shape complementary scores. The molecular interaction was studied using Discovery Studio 3.5 suite.
Results
Screening of miRNAs and genes associated with solid tumor
From the literature, 23 miRNAs were found to be involved in seven solid tumors namely; lung, breast, colorectal, pancreatic, prostate, stomach, and bladder [62]. Again through exploration of online databases 64 different target genes (oncogenes and tumor suppressor genes) were observed to be regulated by these 23 miRNAs (Supplementary Table 1). Again out of these seventeen miRNAs are suspected to be effectively involved in controlling the expression pattern of 45 target genes associated in seven solid tumors (Table 1) [6,7,8,9,10,11,12,13,14,15,16,17,18,19,20,21,22,23,24,25,26,27,28,29,30,31,32,33,34,35,36,37,38,39,40,41]. Further, binding affinity was explored using miRTarbase web server. Fifteen miRNAs and 23 target genes were selected as per the availability of MFE score (Supplementary Table 2) and subjected to further study. The involvement of these 23 genes in different biological functions are explored and inspected from UniProt web server (http://www.uniprot.org/). It was observed only PTEN, TGFBR2, and VEGFA genes mostly regulating angiogenesis, apoptosis, cell cycle, cell proliferation, and other biological functions (Table 2, Fig. 1). Further, the conservation in binding pattern among selected miRNAs and mRNAs (Table 3) were studied through sequence similarity algorithm using Clustal Omega (http://www.ebi.ac.uk/Tools/msa/clustalo/) web server. The study revealed for the existence of quite less portion of conservation at sequence level among chosen 23 mRNAs and also in 15 miRNAs (Fig. 2), is not sufficient to throw light on binding patterns. Therefore, the binding affinity was evaluated between mRNA and miRNA at secondary structural level. Good MFE score suggested, the target genes PTEN, TGFBR2, and VEGFA are having a strong affinity towards miR-106a, miR-21, and miR-29b-2 (Table 3). Hence those genes and miRNAs were taken into consideration to study their interaction at molecular level.
Study of binding affinity between miRNA-mRNA duplex
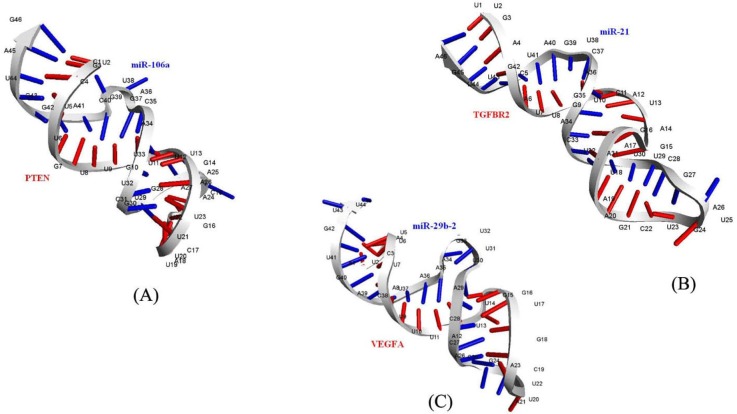
Duplex sequences between miR-106 and PTEN, miR-21 and TGFBR2, miR-29b-2 and VEGFA were extracted from miRTarbase web server and prediction of secondary folding pattern was performed in RNA fold. The secondary folding patterns along with their binding energy value are reported (Table 4). The predicted binding energy suggested for a high affinity among selected miRNA-mRNA duplexes. Predicted three dimensional models [63] of the duplex structures between miR-106 and PTEN, miR-21 and TGFBR2, miR-29b-2 and VEGFA also supported for strong molecular interaction between them.
Structure preparation of AGO protein
The three-dimensional crystallized structure of AGO protein, with 685 amino acid residues was extracted from Protein Data bank (PDB ID: 3F73). The structure of AGO protein contains guide DNA and target RNA duplexes. All water molecules and ligands were removed from the structure. As, the AGO protein is a homo dimer, out of two chains, only chain 'A' of AGO protein was considered and refined before docking. The necessary correction in bond order and bond length of all atoms in the structure was performed using prepare protein and clean geometry protocol of Discovery Studio 3.5.
Study of molecular interaction between miRNAs and AGO protein
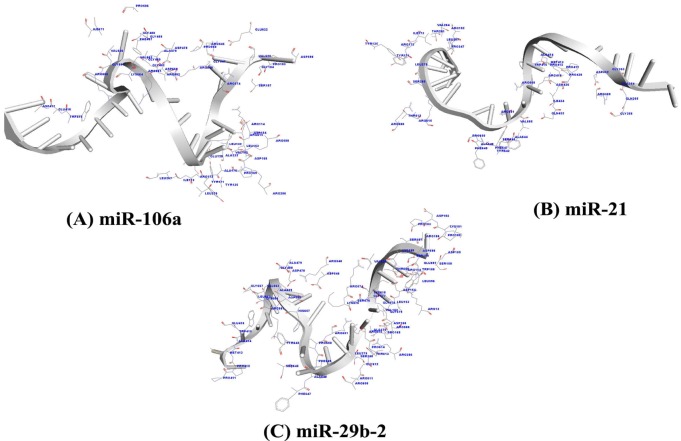
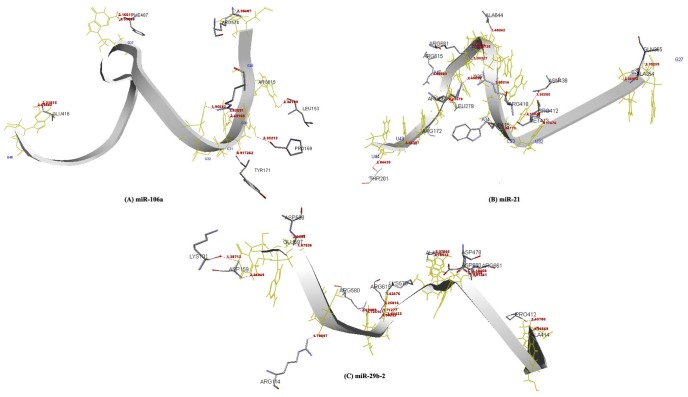
The AGO protein is a key player in the formation of the RNA-induced silencing complex, a major component of RNA interference. The three dimensional structures of miRNA-mRNA duplexes were prepared using RNA COMPOSER web server (Fig. 3). The first round of docking was performed between the miRNAs and AGO (PDB ID: 3F73, chain A) protein to inspect the binding affinity between the complexes. Out of 10 different poses resulted for docking complexes in Patch Dock server, the pose with highest geometrical shape complementary score [64] is considered as the best docked complex. The resulted geometrical shape complementary score and atomic contact energy scores are reported (Table 5), implicated a strong binding affinity between the miRNAs and AGO protein. The binding affinity between the AGO protein and miRNAs (miR-106a, miR-21, and miR-29b-2) is established through the observation of amino acids (miR-106a: LEU 132, ALA 133, VAL 152, LEU 153, ALA 170, ILE 173, LEU 267, LEU 279, ALA 479,VAL 620, VAL 663, VAL 666, and ILE 671; miR-21: ILE 173, VAL 264, LEU 267, LEU 279, ALA 354, ALA 414, ILE 434, ALA 644, ALA 648, and VAL 685; miR-29b-2: VAL 152, LEU 153, ALA 170, LEU 279, ALA 414, ALA 479, LEU 596, VAL 620, ALA 648, LEU 652, ALA 659, LEU 662, and VAL 663) which are strongly hydrophobic in nature and also amino acids with aromatic rings (miR-106a: TYR 135, TYR 171, TRP415, and PHE 487; miR-21: TYR 135, TYR 171, TRP 415, TYR 642, PHE 647, and PHE 649; miR-29b-2: TRP 156, TRP 415, TYR 642, PHE 647, and PHE649) within a distance of 3.5Å (Table 6, Fig. 4). The presence of hydrogen bonding pattern during interaction (Table 7, Fig. 5) also supported the fact of AGO protein driven miRNA based gene regulation.
Study of molecular interaction between AGO protein and miRNA-mRNA duplex
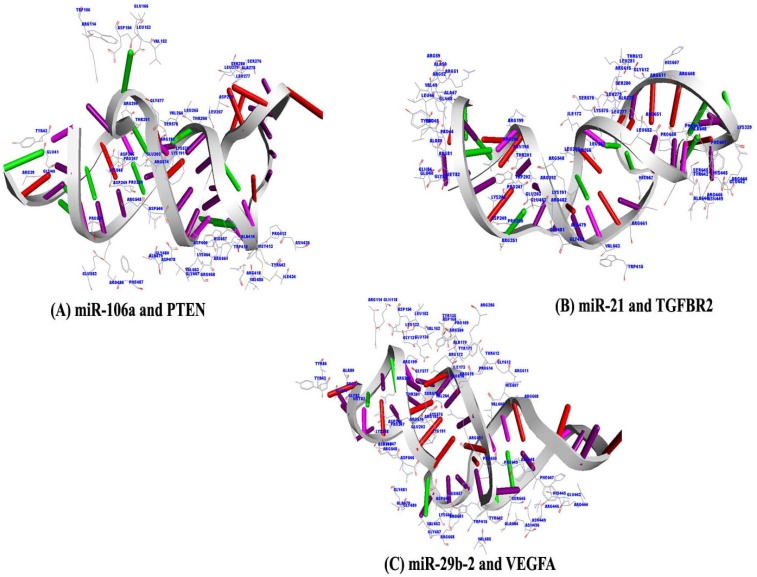
The second round docking was performed between AGO protein and miRNA-mRNA duplexes (miR-106 and PTEN; miR-21 and TGFBR2; and miR-29b-2 and VEGFA) separately in PatchDock web server, and scores are reported (Table 8). Amino acids which are strong hydrophobic in nature and amino acids with aromatic rings which are relatively hydrophobic generally contribute a lot towards the stability of binding during molecular interaction between two macro molecules. Hence the molecular interaction between miRNA-mRNA duplexes and AGO protein was studied by inspecting close amino acid residues of AGO protein within a distant of 3.5 Å. Strong hydrophobic amino acids of AGO like VAL 152, LEU 153, VAL 264, LEU 265, LEU 267, LEU 277, ALA 278, LEU 279, ALA 414, ILE 434, ALA 479, VAL 663, and VAL 685 in miR-106a and PTEN duplex; LEU 45, LEU 46, ALA 47, VAL 49, ALA 50, ALA 80, ILE 173, LEU 265, LEU 267, LEU 277, ALA 278, LEU 279, LEU 281, ALA 479, ALA 644, ALA 648, LEU 652, and VAL 663 in miR-21 and TGFBR2 duplex; and ALA 80, LEU 132, VAL 152, LEU 153, ALA 170, ILE 173, VAL 264, ALA 479, VAL 606, ALA 644, ALA 648, VAL 663, and VAL 685 in miR-29b-2 and VEGFA duplex are observed within a distant of 3.5 Å. Similarly, amino acids like TYR 43, TRP 415, PHE 487, and TYR 642 in miR-106a and PTEN duplex; TYR 43, TRP 202, TRP 415, TYR 642, PHE 647, and PHE 649 in miR-21 and TGFBR2 duplex; and TYR 43, TYR 86, TYR 135, TRP 415, TYR 171, TYR 642, PHE 647, and PHE 649 in miR-29b-2 and VEGFA duplex, with aromatic ring are also found as participating in interaction within the binding pocket of AGO protein around distance of 3.5 Å (Table 9, Fig. 6).
Discussion
The current study focused on in silico identification of genes and their regulation mediated through miRNAs, involved in seven types of solid tumors of colorectal, pancreatic, breast, stomach, lung, prostate, and bladder. miRNAs have a crucial role in gene silencing mechanism observed through the altered expression of certain miRNA molecules in case of neurodegenerative diseases such as Alzheimer and Parkinson suggesting that the association between mRNA-miRNA-AGO, therefore not cancer specific [16,65]. The association between mRNA-miRNA-AGO pattern has been reported [16,65,66] in RNA based gene silencing mechanism and their association at atomic level has already been studied in case of leukemia [25]. In this context, molecular association between AGO protein, miRNAs and mRNAs of target genes were studied and analyzed in case of seven types of solid tumor. Sixty-four genes and 23 miRNAs were collected from the literature (Supplementary Table 1), out of which 45 genes and seventeen miRNAs were selected on the basis of their expression pattern (Table 1). The affinity in binding modes between 15 miRNAs and 23 genes was verified, basing on the availability of MFE scores in miRTarbase web server (Supplementary Table 2). Further, different biological activities of selected genes were studied using UniProt database (http://www.uniprot.org/) to screen out important genes and their contribution towards seven types of solid tumors. The observation suggested three genes namely, PTEN, TGFBR2, and VEGFA were involved in different biological activities like angiogenesis, apoptosis, cell cycle, and cell proliferation, are the major contributing factor in oncogenesis (Table 2). However, quite less sequence conservation was observed within the binding site of 23 target genes (Fig. 2A). But, quite well sequence similarity was observed between miR-106a, miR-17-5p, and miR-20a (Fig. 2B) suggesting for possibility of regulation of PTEN gene through these three miRNAs (Table 3), associated in causing colorectal cancer. Again, a strong and energetically binding affinity was proved through good MFE scores –14.3 (kcal/mol), –16.2 (kcal/mol), and –15 (kcal/mol) for PTEN and miR-106a; TGFBR2 and miR-21; and VEGFA and miR-29b-2, respectively, suggested for appropriate miRNAs selection for PTEN, TGFBR2, and VEGFA genes (Table 3), supported by the predicted binding energy scores for miRNA-mRNA duplex structures (Table 4). Furthermore, to study the molecular basis of AGO protein driven miRNAs namely, miR-106a, miR-21, and miR-29b-2 having highest binding affinity towards their regulating genes namely, PTEN, TGFBR2, and VEGFA respectively, the molecular docking study was carried out. Generally, presence of amino acids with aliphatic group namely, alanine (ALA), valine (VAL), leucine (LEU), and isoleucine (ILE), are strongly hydrophobic in nature provides stability during molecular interaction between macro molecules, whereas amino acids namely, phenylalanine (PHE), tyrosine (TYR), tryptophan (TRP), are relatively hydrophobic in nature, but the presence of aromatic ring like structure provides a steadiness towards binding stability within the structural moiety of a protein. The observation of amino acids (miR-106a: LEU 132, ALA 133, VAL 152, LEU 153, ALA 170, ILE 173, LEU 267, LEU 279, ALA 479,VAL 620, VAL 663, VAL 666, and ILE 671; miR-21: ILE 173, VAL 264, LEU 267, LEU 279, ALA 354, ALA 414, ILE 434, ALA 644, ALA 648, and VAL 685; miR-29b-2: VAL 152, LEU 153, ALA 170, LEU 279, ALA 414, ALA 479, LEU 596, VAL 620, ALA 648, LEU 652, ALA 659, LEU 662, and VAL 663) which are strongly hydrophobic in nature and amino acids with aromatic rings (miR-106a: TYR 135, TYR 171, TRP 415, and PHE 487; miR-21: TYR 135, TYR 171, TRP 415, TYR 642, PHE 647, and PHE 649; miR-29b-2: TRP 156, TRP 415, TYR 642, PHE 647, and PHE649) during interaction within a distance of 3.5Å (Table 6, Fig. 4) recognized a strong molecular interaction between AGO protein and miR-106a, miR-21, and miR-29b-2. Again, weak interaction like hydrogen bonding has a major contribution to facilitate the stability of molecules during interaction at atomic level. Here, the amino acids like, LEU 153, PRO 169, TYR 171, GLU 416, PHE 487, ARG 574, and ARG 615; ARG 172, THR 201, LEU 279, ALA 354, GLN355, PRO 412, MET 413, TRP 415, ARG 418, ASN 436, ARG 615, ALA 644, ARG 661, and ARG 668; LYS 101, ARG 114, ASP 159, PRO 412, ALA 414, ASP 478, LYS 575, ARG 580, ARG 580, GLU 597, ASP 598, ARG 615, ALA 648, ASP 660, and ARG 661 are participated in hydrogen bonding interaction with miR-106a, miR-21, and miR-29b-2 respectively in the structural moiety of AGO protein within a distance of 2.5Å (Table 7, Fig. 5), strongly supports for binding stability during assistance of AGO protein for miRNA based gene regulation. Furthermore, presence of commonly participating strong hydrophobic amino acids namely, VAL 663 and amino acid with aromatic rings namely, TYR 43, TRP 415, and TYR 642 during molecular interaction of AGO protein with miR-106a and PTEN, miR-21 and TGFBR2, and miR-29b-2 and VEGFA duplex within a distance of 3.5Å (Table 9, Fig. 6). Although, the evidence at molecular association between mRNA-miRNA-AGO pattern have already been established computationally [16,61] the current investigation recommending for AGO protein assistance in regulation of PTEN, TGFBR2, and VEGFA genes by miR-106a, miR-21, and miR-29b-2, respectively, associated in seven types of solid tumor. This report would be helpful in understanding miRNA-based gene silencing mechanisms in seven types of solid cancer like colorectal cancer, pancreatic cancer, breast cancer, stomach cancer, lung cancer, prostate cancer, and bladder cancer. Furthermore, extensive computational study may be carried out for better understanding the mechanism of miRNAs based gene regulation in solid cancer. This methodology would further help us to design suitable miRNAs against respective genes rationally, only after validation through laboratory experiments.